


A study reveals that a process called loop extrusion, which is central to the folding and function of chromosomes, also seems to play a key part in the repair of double-strand DNA breaks.
One of the challenges that all organisms face is how to compress long DNA molecules — two metres long in the case of human DNA — into the cell nucleus in an ordered and tidy fashion. The DNA must also be protected from damage. Writing in Nature, Arnould et al.1 highlight the ingenuity with which cells achieve these aims: one of the mechanisms involved in packing and folding DNA, namely loop extrusion, is also involved in repairing damage.
Cells neatly compact their spaghetti of DNA in several ways. In cells with a nucleus, the DNA is first wrapped around cores of histone proteins to make structures called nucleosomes, which together form a chromatin fibre that looks like beads on a string. Loop extrusion is the subsequent compaction process, whereby a molecular motor binds a chromatin fibre and reels it in from the sides, forcing out a progressively larger loop in between (Fig. 1a).
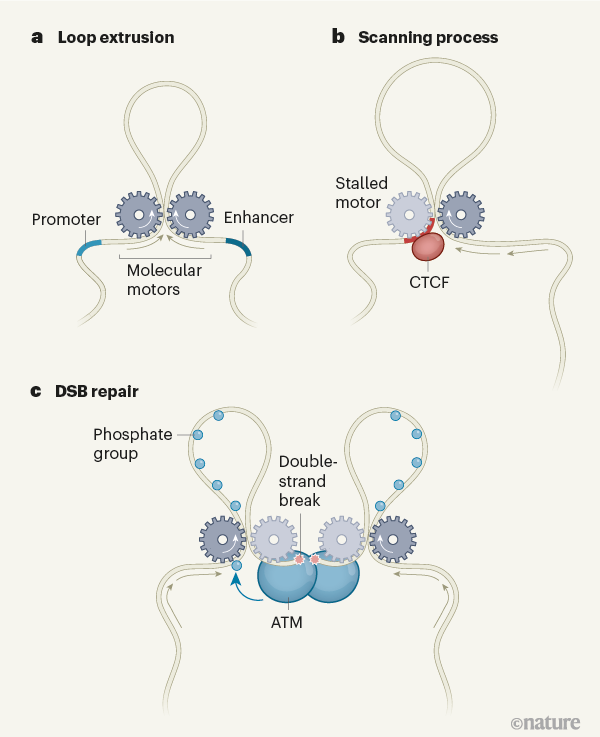
Although the process of loop extrusion was hypothesized decades ago, only in the past few years has it become clear that it is a universal mechanism that organizes DNA in organisms from bacteria to humans. In 2016, computational models showed that extrusion can compact DNA, turning a hairball of chromatin into detangled yet tightly packed chromosomes. Simulations also indicated that, when extrusion is stalled by barriers on the chromatin (a normal part of the process), it produces chromosomal domains seen in data from Hi-C — a technique used to characterize chromosome structure.
These studies further suggested that ‘structural maintenance of chromosomes’ (SMC) complexes — once thought to be passive rings or staples — are actually loop-extruding motors. Moreover, proteins known as CCCTC-binding factors (CTCFs), which attach to specific DNA sequences, were proposed to be barriers that catch and stall SMC motors called cohesins (Fig. 1b). All in all, a range of experimental evidence — from in vivo depletion of SMCs and CTCFs, to direct visualization of single molecules — now supports the existence of loop extrusion.
Cellular processes often multitask. So, could this ubiquitous mechanism for weaving the genome have other functions? During cell division, the formation of loops is key to compacting chromosomes to enable accurate passing of genetic material into daughter cells. But the role of extrusion during interphase — the period during which cells duplicate their DNA and grow in preparation for the next division — is yet to be understood.
There are several possibilities. One of these is in regulating gene expression: extrusion can bring together distant genomic elements (such as enhancers and promoters, which regulate transcription) that are not separated by a CTCF barrier. Such barriers can also turn extrusion into genomic tracking. To explain, when a cohesin protein stalls on one CTCF, it can continue reeling DNA in on the other side, thereby tracking over long genomic regions (Fig. 1b). This mechanism has been implicated in the stochastic expression of certain cell-adhesion proteins and the random rearrangement of antibody gene segments, resulting in striking antibody diversity. Arnould et al. add to this list of possibilities, suggesting that loop extrusion safeguards the genome by supporting the repair of double-strand DNA breaks (DSBs).
To repair severed DNA, cells must first establish a large (roughly one million base pair) region of modified H2AX histones flanking the break. Phosphate groups are added to the histones by certain enzymes, including one called ATM, producing a region of what are known as γH2AX histones. These histones signal the presence of the break to repair enzymes.
Arnould et al. confirm observations that γH2AX regions closely resemble chromosomal domains that are established by loop extrusion and demarcated by CTCFs. They also find that the ATM that establishes the modifications remains bound at the DSB (Fig. 1c). How, then, can this ATM reach histones located a million base pairs away? The group provides evidence that genome folding is key. The asymmetry of the modified regions relative to the DSB, with limited spreading of histone modifications beyond CTCF barriers, indicates that the spreading is mediated by a ‘directional’ mechanism that tracks along the genome and stops at CTCFs, rather than by spatial contacts made between disparate chromosome regions or a phase separation that would establish contact symmetrically and wouldn’t obey CTCF barriers. Extrusion readily provides such a directional mechanism.
Furthermore, the temporal dynamics of γH2AX spreading support a directional process. A previous study showed that the modifications established by ATM spread directionally, rather than by spatial contacts or the more random process of diffusion. Another study found that γH2AX spreads directionally at a rate of 150,000 bases per minute. This speed is consistent with the speed of loop extrusion measured in vitro and in bacteria.
One possibility for the directional spreading of γH2AX, then, is that DSB-bound ATM uses loop-extruding SMCs on either side of the break to scan flanking DNA. To achieve this, a break or ATM would stall each SMC on one side, similar to CTCF, while still allowing it to reel chromatin in on the other side, progressively pulling more-distant regions to be modified by the stationary ATM. Such stalling should generate a characteristic pattern of stripes on a Hi-C map — as Arnould et al. and others found.
The evidence that cohesin itself is the primary loop extruder remains modest, however. Arnould and colleagues found that loss of one cohesin subunit (Scc1) led to loss of extrusion stripes near DSBs on Hi-C maps, but that depleting or stabilizing cohesin itself had only slight effects on γH2AX profiles. It’s possible that other SMC complexes, such as SMC5/6 or MRX, or an Scc1-independent cohesin have the predominant role here. Data from another study hint at this: extrusion stripes occur near DSBs in dividing yeast cells, but diminish following depletion of MRX (rather than cohesin). There might well be a complex web of loop-extrusion activities, mediated by different motors.
The findings could have implications well beyond DNA repair. Histone modifications are key to development and cellular identity, but the mechanisms that establish and maintain them are poorly understood. The suggestion of a role for the SMC–CTCF system in spreading histone modifications s radically different from the known mechanism underlying this process. Until now, extrusion- and modification-dependent folding mechanisms were considered to be entirely separate.
It is also possible that the roles of cohesin and other SMCs in repairing breaks go beyond γH2AX spreading. Cohesin has long been implicated in DNA repair, but was thought to be a passive ring, keeping together duplicated chromosomes for repair. The findings suggest more-active roles, for example in enabling the search for a region of DNA that can act as a template for repair. What’s more, the recruitment of SMCs to breaks, and the central role of cohesin in other processes that deal with DSBs (such as a type of division called meiosis and immunoglobulin-gene regulation), suggests that these motors might be important in detecting and managing broken DNA during many normal cellular processes. So, SMC complexes — master weavers of the genome — might be in charge not only of making loops, but also of finding and tying together broken fibres.
Nature 590, 554-555 (2021)





